1.2 Строение микроскопических грибов. Есть клеточная стенка у грибов
Химический состав и структура клеточной стенки грибов
Клеточная стенка грибов многослойная, при этом разные слои образованы различающимися по химическому составу структурными углеводами, которые по химическому составу можно разделить на 3 группы:
полимеры глюкозы (глюкан, хитин, целлюлоза). Глюканы составляют наружный слой клеточной стенки большинства грибов. Внутренний слой грибной клеточной стенки образован цепочками хитина, придавая ей жесткость. Хитин замещает целлюлозу, которая у большинства грибов отсутствует, но входит в состав клеточной стенки оомицетов, которые в настоящее время к типичным грибам не относятся. Деацетилированный хитин получил название хитозан, который в комплексе с хитином образует клеточную стенку зигомицетов.
полимеры других моносахаридов (маннозы, галактозы и др.) в отличие от высших растений, где они составляют основу матрикса под общим названием гемицеллюлоза, менее характерны для грибов. Исключение составляют дрожжи, в клеточных стенках которых особенно много полимеров маннозы, называемых маннаны. Предполагают, что такой состав стенки лучше обеспечивает почкование.
полимеры углеводов, ковалентно связанные с пептидами (гликопротеины) формируют срединный слой многослойной клеточной стенки и играют важную роль, как в поддержании структурной целостности клеток, так и в ее обменных процессах с окружающей средой.
К другим специфическим особенностям грибной клетки относятся: отсутствие пластид, что сближает ее с клеткой животных;
Отсутствие крахмала, который у эумицетов замещается полисахаридом, близким к животному крахмалу гликогену, у оомицетов – полисахаридом, близкому к ламинарину бурых водорослей. Вырабатывается и целый ряд специфических для грибов запасных углеводов.
Выработка специфических вторичных метаболитов, из которых большую роль играют антибиотики, фито– и микотоксины, фитогормоны.
К специфическим особенностям грибов относятся также гетерокариоз и парасексуальный процесс.
У грибов очень широко распространено явление гетерокариоза или разноядерности, при котором в одной клетке длительное время сохраняются ядра, гетероаллельные по некоторым генам. Гетерокариоз заменяет гаплоидным грибам гетерозиготность и способствует быстрой адаптации грибов к меняющимся условиям. Наличие разноядерности обусловлено рядом специфических особенностей грибов:
1. наличие более одного ядра в клетке
2. специфическое строение клеточной перегородки, в которой имеется одно или несколько сквозных отверстий, называемых порами, через которые ядра могут мигрировать из одной клетки в другую
3. гифы внутри одной колонии и даже разных близко расположенных колоний, выросших из разных спор одного вида гриба часто срастаются, в результате чего возможен обмен ядрами разных штаммов.
Парасексуальный (псевдополовой) процесс. Если в гетерокариотичных клетках сливаются ядра, гетероаллельные по какому-либо локусу, возникает гетерозиготное диплоидное ядро. Оно может попасть в спору и дать начало диплоидному гетерозиготному клону. В процессе митозов диплоидные ядра могут возвращаться в гаплоидное состояние вследствие потери одного набора хромосом или в них может происходить обмен участками хромосом (митотический кроссинговер). Оба процесса сопровождаются рекомбинацией родительских генов и, следовательно, фенотипов. Парасексуальная (бесполовая) рекомбинация очень редкое явление, не превышающее одного ядра на миллион, но вследствие огромного числа ядер в мицелии постоянно наблюдается в популяциях грибов.
Размножение – вегетативное, бесполое, половое.
Вегетативное – фрагментация таллома, образование хламидоспор, которые после периода покоя прорастают в мицелий, почкование у дрожжей.
Бесполое размножение у разных грибов может осуществляться подвижными и неподвижными спорами. Зооспоры образуют относительно небольшая группа грибов и грибоподобных организмов – водные и некоторые наземные, у которых отчетливо прослеживаются генетические связи с водными грибами и водорослями. Структура жгутиков является важным диагностическим признаком при отнесении к определенному царству. У подавляющего большинства грибов при бесполом размножении образуются неподвижные споры, что указывает на их очень давний выход на сушу. По месту формирования и локализации различают эндогенные, образующиеся в спорангиях спорангиоспоры и экзогенные (конидии), развивающиеся на специальных гифах – конидиеносцах. Конидии образуются у большинства грибов (аскомицеты, базидиомицеты, некоторые зигомицеты) составляя конидиальное спороношение, которое отличается огромным многообразием и широко используется для диагностики грибов.
Половое размножение грибов имеет свою специфику как в морфологии полового процесс, так и в механизмах генетической и физиологической регуляции пола и передачи наследственной информации.
Соматогамия – самый распространенный и наиболее простой тип полового процесса, заключается в слиянии двух не дифференцированных на гаметы соматических клеток. Иногда он протекает даже без слияния клеток – сливаются ядра внутри клетки. Встречается у большинства базидиомицетов, сумчатых дрожжей и некоторых др. таксонов.
Гаметангиогамия – на гаплоидном мицелии обособляются гаметангии, при половом процессе сливается их содержимое. Такой половой процесс характерен для большинства сумчатых грибов. Вариантом гаметангиогамии является зигогамия у зигомицетов.
Гаметогамия в виде изо- гетеро и оогамии у грибов встречается значительно реже, чем у других эукариот. Изо- и гетерогамия встречается лишь у хитридиомицетов. Классическая оогамия с образованием сперматозоидов и яйцеклеток у грибов не выражена, а встречаются сильно измененные варианты.
По особенностям регуляции пола у грибов выделяют несколько типов полового процесса
Гинандромиксис можно рассмотреть на примере двудомных оомицетов, у которых оогонии и антеридии развиваются на разных талломах, например фитофтора или картофельный гриб. Если генетически однородный штамм выращивать в монокультуре, то он размножается только бесполым способом. Если рядом оказываются мицелии двух штаммов, то у них прослеживаются морфогенетические изменения под действием химических выделений стероидной природы – половых феромонов. Антеридиол индуцирует образование антеридиев у партнера, а оогониол –оогониев. При этом регуляция пола имеет относительный характер: будет ли штамм образовывать антеридии или же оогонии, зависит от количественного соотношения соответствующих феромонов у него и его партнера. Отсюда название полового процесса – гинандромиксис.
Димиксис или гетероталлизм. Уже давно было подмечено, что грибы могут быть гомо- или гетероталличными. У гомоталличных видов при половом процессе сливаются генетически идентичные ядра внутри мицелия. У гетероталличных видов для прохождения полового цикла необходимо на каком-то этапе (различном у разных видов грибов) слияние потомков спор (точнее, их ядер). Для половой совместимости двух штаммов необходимо генетическое различие (гетероаллельность) в определенных локусах, называемых локусами спаривания. У большинства грибов (зигомицеты, аскомицеты, часть базидиомицетов) имеется один локус спаривания с двумя аллелями. Локус спаривания состоит из нескольких генов, котрые управляют синтезом половых феромонов. Такой гетероталлизм называется однофакторным или биполярным. Потомство таких грибов после мейоза разделяется на две самонесовместимые, но взаимно совместимые группы в соотношении 1:1, т.е. вероятность родственного(инбридинга) и неродственного(аутбридинга) скрещивания составляет как и у двуполых высших эукариот 50%.
В геноме высших базидиальных грибов имеется два локуса спаривания – A и B, причем совместимы друг с другом только штаммы гетероаллельные по обоим локусам (Ax Bx совместим с Ay By, но не с Ax By и Ay Bx). Такой гетероталлизм называют двухфакторным или тетраполярным. Он снижает вероятность инбридинга до 25 %.
Диафоромиксис – у высших базидиомицетов имеется не два, а много аллелей локуса спаривания, случайно встречающихся у разных штаммов, слагающих популяцию. Такой контроль спаривания обеспечивает 100% вероятность аутбридинга, так как штаммы, имеющие разные аллели, взаимносовместимы, а аллелей очень много. В результате образуются панмиксные гибридные популляции.
Жизненные циклы грибов также разнообразны как и сами грибы. Основные циклы, их принадлежность к отделам грибов
1.Бесполый цикл характерен для огромной группы несовершенных или митогрибов, утративших половое размножение. Деление их ядер исключительно митотические. Подавляющее большинство митогрибов относится к сумчатым грибам, но в связи с утратой полового процесса, составляют формальную группу несовершенных грибов или дейтеромицетов.
2. Гаплоидный цикл. Вегетативный таллом несет гаплоидные ядра. После полового процесса (сингамии) диплоидная зигота (обычно после периода покоя) делится мейотически – зиготический мейоз. Характерен для зигомицетов и многих хитридиомицетов.
3. Гаплоидно-дикариотический цикл характеризуется тем, что после слияния содержимого гаметангиев (гаметангиогамия) или соматических клеток гаплоидного мицелия (соматогамия) ядра образуют дикарионы (пары генетически разных ядер). Они синхронно делятся, образуя дикариотический мицелий. Половой процесс завершается слиянием ядер дикариона, образующаяся зигота делится мейозом без периода покоя. Мейоспоры составляют половое спороношение сумчатых и базидиальных грибов в виде аскоспор и базидиоспор. При их прорастании образуется гаплоидный мицелий. У подавляющего большинства сумчатых грибов (кроме дрожжей и тафриновых грибов) в жизненном цикле преобладает гаплоидная фаза в виде вегетативного мицелия (анаморфа), дикариотическая фаза кратковременна и представлена аскогенными гифами, на которых образуются сумки (телеоморфа). У базидиальных грибов в жизненном цикле преобладает дикариотическая фаза, гаплоидная фаза кратковременна.
4. Гаплоидно-диплоидный цикл в виде изоморфной смены поколений у грибов встречается редко (некоторые дрожжи и водные хитридиомицеты).
5. Диплоидный цикл характерен для оомицетов и некоторых сумчатых дрожжей. Вегетативный таллом диплоидный, мейоз гаметический, наблюдается при формировании гаметангиев или гамет.
3. Экологические группы грибов.
Грибы и грибоподобные организмы входят во все наземные и водные экосистемы, как важнейшая часть гетеротрофного блока, вместе с бактериями занимая трофический уровень редуцентов. Широкое распространение грибов в биосфере определяется рядом важнейших особенностей:
1. Наличие у большинства мицелиальной структуры таллома. (позволяет быстро осваивать субстрат, иметь большую поверхность контакта со средой).
2. Большая скорость роста и размножения, позволяющие в короткие сроки заселять большие массы субстрата, образуя огромное число спор и распространять их на большие расстояния.
3. Высокая метаболическая активность, проявляющаяся в широком диапазоне значений экологических факторов.
4. Высокая скорость генетической рекомбинации, значительная биохимическая изменчивость, экологическая пластичность.
5. Способность быстро переходить к состоянию анабиоза, переживать неблагоприятные условия в течение длительного времени.
Главным экологическим фактором для грибов является пищевой субстрат. По отношению к этому фактору выделяют основные группы грибов, которые называются трофическими группами.
1. Сапротрофы – обитают на растительных остатках
4. Краткая характеристика отделов грибов.
Похожие статьи:
poznayka.org
Клеточная стенка | Биология
В отличие от животных и многих простейших, у растений, бактерий и грибов, почти все клетки имеют стенку, лежащую кнаружи от цитоплазматической мембраны и обладающую повышенной прочностью. Основная функция данной структуры — опора и защита.
Клеточные стенки (или клеточные оболочки) строятся из веществ, синтезируемых самими клетками. Их химический состав различен у растений, грибов и прокариот. Кроме того, даже у одного растения у различных клеток состав стенок несколько различен.
Клеточная стенка растений состоит в основном из целлюлозы. Целлюлоза — это полисахарид, мономером которого является глюкоза.
Основу бактериальных клеточных стенок составляет вещество муреин (относится к пептидогликанам). У грамположительных бактерий в состав оболочки входят различные кислоты, а сама оболочка плотно прилегает к цитоплазматической мембране. У грамотрицательных бактерий оболочка более тонкая и не прилегает к мембране. Между мембраной и оболочкой образуется периплазматическое пространство. Снаружи клеточная оболочка грамотрицательных прокариот окружена внешней мембраной, составленной из липополисахарида.
У грибов основным веществом клеточных стенок является хитин, а не целлюлоза.
Состав клеточной стенки растений
У растений стенка дочерних клеток образуется уже во время деления родительской. Впоследствии она называется первичной. У многих клеток позже образуется вторичная оболочка.
Первичная клеточная оболочка состоит из микрофибрилл целлюлозы, погруженных в матрикс из других полисахаридов. Отличительной особенностью волокон целлюлозы является их прочность. Молекула целлюлозы представляет собой длинную полисахаридную цепь. Отдельные молекулы соединяются друг с другом водородными связями в пучок, который называется микрофибриллой. Такие фибриллы образуют каркас клеточной стенки.
Матрикс клеточной стенки составляют полисахариды пектины и гемицеллюлозы, а также ряд других веществ (например, белков). Пектиновые вещества представляют собой группу кислых полисахаридов, их молекулы могут быть не только линейными, но и разветвленными. Гемицеллюлозы также смешанная группа полисахаридов. Длина их линейных молекул короче, чем у целлюлозы.
Оболочки соседних клеток растений соединены между собой срединной пластинкой, состоящих из пектатов магния и кальция, для которых характерна клейкость.
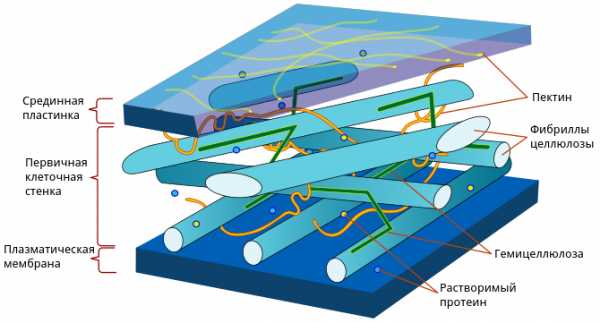
В состав стенок растений входит вода (составляет более половины массы), обуславливая ряд физических и химических свойств полисахаридов.
Жесткий каркас растения во многих местах пронизан каналами (плазмодесмами), по которым цитоплазма одной клетки соединяется с цитоплазмой соседних.
Клетки мезофилла листа (а также некоторые другие) на протяжении всей своей жизни имеют только первичную стенку. У большинства же клеток на первичную оболочку с внутренней стороны отлагается вторичная стенка, составленная из дополнительных слоев целлюлозы. Обычно в это время клетка уже дифференцирована и не растет (исключение составляют, например, клетки колленхимы).
В каждом отдельном слое вторичного утолщения микрофибриллы целлюлозы располагаются под одним углом (параллельно друг другу). Однако разные слои имеют разный угол, что обеспечивает большую прочность.
Часть клеток растений одревесневают (трахеальные элементы ксилемы, склеренхима и др.). В основе этого процесса лежит интенсивная лигнификация стенок (в небольших количествах лигнин есть во всех оболочках). Лигнин не является полисахаридом, а представляет собой сложное полимерное вещество. Отложения лигнина могут иметь различную форму (сплошную, кольцевую, спиральную, сетчатую). Он скрепляет целлюлозу, не дает ей смещаться. Лигнин не только обеспечивает прочность, но и дает дополнительную защиту от неблагоприятных физических и химических факторов.
Функции клеточной стенки
Оболочки разных клеток совместно обеспечивают всему растению и его отдельным частям механическую прочность и опору. Это функция клеточной стенки аналогична одной из функций скелета животных. Однако она не единственная.
Жесткость стенок препятствует растяжению клеток и их разрыву. В результате по физическим законам в клетки может путем осмоса поступать вода. Для травянистых растений тургоцентричность клеток является единственной их опорой.
Микрофибриллы целлюлозы ограничивают рост клеток и определяют их форму. Если микрофибриллы окольцовывают клетку, то она будет расти в длину (поперек направления волокон).
Связанные клеточные стенки образуют апопласт, по которому передвигается вода и минеральные вещества. Плазмодесмы связывают содержимое разных клеток в единую систему — симпласт.
Стенки сосудов ксилемы, трахеид, ситовидных трубок выполняют транспортную функцию.
Наружные клеточные стенки эпидермальных клеток покрыты воском (кутикулой). С одной стороны, он препятствует испарению воды, с другой – проникновению вредных микроорганизмов.
У некоторых растений в определенных клетках оболочки видоизменяются и служат местом запаса питательных веществ.
biology.su
Клеточная стенка | Info-Farm.RU
Клеточная стенка — достаточно твердый слой, окружающий клетку. Она располагается за пределами цитоплазматической мембраны (также известной в некоторых случаях, как клеточная мембрана) и обеспечивает дополнительную поддержку и защиту. Клеточные стенки найдены у бактерий, архей, грибов и растений
Если разрушить клеточную стенку с помощью некоторых ферментов, то останется, то есть клетка окружена клеточной мембраной, называется протопластом. В отличие от мембран, клеточная стенка не в состоянии контролировать транспорт молекул в клетку из внешней среды.
Клеточные стенки высших растений
Растительные клеточные стенки выполняют целый ряд функций: они обеспечивают жесткость клетки для структурной и механической поддержки, придают форму клетке, направление ее роста и в конечном счете морфологию всему растению. Клеточная стенка также противодействует тургора, то есть осмотическому давлению, когда дополнительное количество воды поступает в растения. Клеточные стенки защищают против патогенов, проникающих из окружающей среды, и запасают углеводы для растения. Растительные клеточные стенки строятся прежде всего с углеводного полимера целлюлозы.
В течение цитокинеза сначала с клеточной пластинки формируется промежуточная ламела, и первичная клеточная стенка расширяется внутри промежуточной ламели. Фактическая структура клеточной стенки не всегда четко определена, существуют несколько моделей ее структуры: модель перекрестных ковелентних связей, модель привязи, модель диффузного слоя и модель стратифицированного слоя. Однако, первичная клеточная стенка может быть определена как сеть волокон целлюлозы направленных в случайных направлениях. Волокна воздержатся вместе водородными связями, обеспечивает достаточно высокую прочность. Клетки держатся вместе и разделяют желатиновую мембрану, называет его промежуточная ламела, которая содержит пектаты (соли пектиновой кислоты) магния и кальция. Клетки взаимодействуют через плазмодесмы, то есть цитоплазмени канальцы, соединяющие цитоплазмы клеток через клеточные стенки.
В некоторых типах клеток некоторых растений, после того как достигается максимальный размер или определенная точка развития, между растительным клеткой и первичной стенкой формируется вторичная стенка. В отличие от первичной стенки, волокна выравниваются в основном в одном направлении, и с каждым дополнительным слоем их ориентация слегка меняется. Клетки с вторичными клеточными стенками очень жесткие. Межклеточное взаимодействие все еще возможна через отверстия во вторичной клеточной стенке, позволяющие плазмодкрми проникать через обе клеточные стенки.
Основные углеводы, составляющих первичную клеточную стенку, это целлюлоза, гемицеллюлоза и пектин. Целлюлозные микроволокна связываются через хемицелюлозни мостики, формируя целлюлозно-хемицелюлозну сеть, которая окружена матрицей пектина. Самый общий тип хемицелюлозы в первичной клеточной стенке — ксилоглюкан.
Растительные клеточные стенки также содержат ряд белков, распространенные включают богатые гидропролин гликопротеины (HRGP), также известные как екстенсины, арабиногалактан протеины (AGP), богатые глицин протеины (GRP) и богатые пролин протеины (PRP). За исключением богатых глицин протеинов, все эти белки гликозилированные и содержат гидроксипролин (Hyp). Каждый класс гликопротеинов определяется характерной, часто повторяющейся последовательности аминокислот. Причудливые протеины содержат два или более областей, каждая из последовательностью от другого класса гликопротеинов. Большинство белков клеточной стенки перекрестных связаны с углеводами клеточной стенки и, возможно, имеют структурные функции.
Вторичные клеточные стенки могут содержать лигнин и суберин, делая стенки твердыми. Относительный состав углеводов, дополнительных компонентов и белков зависит от вида растения, типа и возраста клеток.
Клеточные стенки водорослей
Подобно высших растений, большинство водорослей, независимо от происхождения, имеют клеточные стенки. Клеточные стенки водорослей содержат целлюлозу и многие другие гликопротеинов. Дополнительные полисахариды в клеточных стенках водорослей раньше часто использовали как один из признаков для установления их таксономии, но некоторые несвязанные группы имеют те же полисахариды.
- Манозилови микроволокна зучтричаються в клеточных стенках многих зеленых водорослей (примитивних растений), в том числе родов Codium, Dasycladus и Acetabularia, и некоторых красных водорослей например, родов Porphyra и Bangia.
- Ксилан
- Альгиновая кислота — обычный полисахарид в клеточных стенках бурых водорослей
- Сульфоновани ролисахариды встречаются клеточных стенках большинства водорослей, например, в красных водорослях находим агарозу, каррагинан, порфиран, фурселеран и фуноран.
Другие вещества, которые могут быть найдены в водорослевых клеточных стенках, включают спорополенин и кальций.
Группа диатомовых водорослей (диатомей) синтезируют клеточные стенки (также известно как фрустулы или клапаны) с Кремнев кислоты (особенно ортокремневои кислоты, H 4 SiO 4). Кислота полимеризуется внутриклеточно, затем стенка вытесняется наружу для защиты клетки. Такие стенки требуют меньше энергии на свой синтез, позволяя сохранить много энергии, что, возможно, частично отвечает за быстрый рост этих организмов.
Клеточные стенки грибов
Не все виды грибов имеют клеточные стенки, но если стенка присутствует, она состоит из глюкозамина и хитина, того же углерода, что придает прочность экзоскелета насекомых. Они служат подобной цели, что и клеточные стенки растений, предоставляя грибным клеткам твердость и поддерживать свою форму и предотвращения осмотическому лизису. Они также ограничивают вход молекул, потенциально ядовитых для гриба, например некоторым фунгициды растительного и искусственного происхождения. Состав, свойства и форма грибной клеточной стенки меняются на протяжении жизненного цикла и зависят от условий произрастания.
Другие эукариоты
Оомицеты, патогены растений похожи на грибы, также имеют целлюлозные клеточные стенки. До недавнего времени за этот признак их относили к грибам, структурные и молекулярные свидетельства заставили отнести их к гетероконтив, как и бурые водоросли.
Клеточные стенки бактерий
Как в других организмах, бактериальные клеточные стенки обеспечивают структурную целостность клеткам. У прокариот, первичная функция клеточной стенки — защитить ячейку от внутреннего тургора, вызванным высокой концентрацией белков и инших молекулы внутри клетки по сравнению с окружающей средой. Бактериальная клеточная стенка отличается от других тем, что ее основным компонентом является пептидогликан, слой которого размещается немедленно вне цитоплазматической мембраны. Пептидогликан отвечает за твердость стенки и придания формы клетке. Он относительно пористый и не мешает потока растворимых молекул сквозь него. Существует два основных типа бактериальных клеточных стенок, по этому признаку бактерии делятся на грам-отрицательные и грам-положительные.
Грам-положительные бактерии
Грам-положительные бактерии характеризуется присутствием очень толстого слоя пептидогликана, который отвечает за содержание красителя кристал-виолет течение процедуры окрашивания по Граму. Такая стенка находится исключительно в организмах, принадлежащих к типам Actinobacteria (или грамм-положительные организмы с высоким содержанием G + C) и Firmicutes (или грамм-положительные организмы с низким содержанием G + C). Бактерии в пределах типа Deinococcus-Thermus также могут положительно краситься по Граму, но содержат некоторые структуры клеточной стенки, типичные от грамотрицательных организмов. В клеточную стенку грамположительных бактерий встроенные полимерного спирты, тейхоевая кислота, некоторые из которых связываются с липидами, формируя липотейхоеву кислоту. Ци вещества отвечают за соединение пептидогликана с цитоплазматической мембраной. Тейхоевая кислота придает клетке отрицательный электрический заряд благодаря наличию фосфодиестерних связей между мономерами тейхоевая кислоты.
Грамотрицательные бактерии
В отличие от грамположительных бактерий, грамотрицательные бактерии содержат очень тонкий слой пептидогликана, отвечающий за неспособность клеточных стенок содержать краситель кристал-виолет течение процедуры окрашивания по Граму. В дополнение к слою пептидогликанов, грамотрицательные бактерии имеют вторую, так называемую внешнюю мембрану, находится кнаружи от клеточной стенки и компонует фосфолипиды и липополисахариды на своей внешней стороне. Отрицательно заряженные липополисахариды также предоставляют клетке отрицательный электрический заряд. Химическая структура липополисахаридив внешней мембраны часто уникальная для отдельных штаммов бактерий и часто отвечает за реакцию антигенов с представителями этих штаммов.
Как любой двойной слой фосфолипидов, внешняя мембрана достаточно непроницаема для всех заряженных молекул. Однако, белковые каналы (погрузится) присутствуют во внешней мембране, позволяют пассивный транспорт многих ионов, сахара и аминокислот через внешнюю мембрану. Таким образом, эти молекулы присутствуют в периплазматическое, слое между внешней и цитоплазматической мембранами. Периплазматическое содержит слой пептидогликана и много белков, шо отвечают за гидролиз и прием внеклеточных сигналов. Читается, что перивлазма гелеобразная, а не жидкая, из-за высокого содержания белка и пептидогликана. Сигналы и живильни вещества с периплазматическое попадают в цитоплазму клетки используя транспортные белки в цитоплизматичний мембране.
Клеточные стенки архей
Хотя они и не уникальны, клеточные стенки архей несколько отличаются от бактериальных. Например, клеточные стенки большинства архей образованные внешними слоями белков или S-слоем. S-слоя распространенные в бактериях, где они служат единственным компонентом клеточной стенки в некоторых организмах (например в Planctomyces) или внешним слоем во многих организмах с пептидогликана. За исключением одной группы метаногенов, археи не имеют пептидогликаннои стенки. Даже в данном случае, пептидогликаны очень отличается от типа, найденного в бактериях.
Изображения по теме

info-farm.ru
Строение и функции клеточной стенки — Науколандия
Клеточная стенка — это дополнительная оболочка, которая располагается поверх (с внешней стороны) цитоплазматической мембраны и образуется в процессе жизнедеятельности самой клеткой. Такая оболочка есть у клеток не всех организмов, а только у растений, грибов, бактерий, части простейших (одноклеточных эукариот). Ее нет у животных клеток и многих простейших.
Строение и функции клеточной стенки взаимосвязанно формировались в процессе эволюции. При этом ее химическое строение (в большей степени) и функции (в меньшей) у разных групп организмов различаются. Так у растений основным компонентом оболочки является целлюлоза, у грибов — хитин, у бактерий — муреин.
Обычно в школьном курсе цитологии подробно рассматриваются строение и функции растительной клеточной стенки (оболочки).
Целлюлоза представляет собой линейный полисахарид, мономером которого является глюкоза. В составе клеточной стенки молекулы целлюлозы соединяются между собой водородными связями и образуют микрофибриллу (пучок). В оболочке множество таких фибрилл. Часть из них расположены параллельно друг другу, другая часть — под углом к первой и т. д. Такое строение создает прочный каркас.
Кроме целлюлозы, в состав клеточной стенки растений входят другие вещества (вода, гемицеллюлоза, пектиновые вещества, белки и др.). Они формируют матрикс, в котором находятся фибриллы. Вода составляет 60-70% массы оболочки. Молекулы гемицеллюлозы более короткие и разветвленные по-сравнению с целлюлозой, они связывают между собой микрофибриллы.
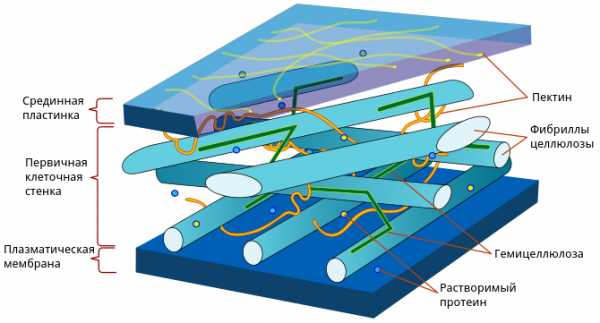
Пектины также представляют собой полисахариды (линейные и разветвленные), основным мономером которых является галактуроновая кислота. Также в их состав входят арабинозы и галактозы, остатки метанола. Пектиновые вещества имеют кислую природу, могут быть растворимыми и нерастворимыми. Растворимые пектины при добавлении сахара переходят в гелеобразное состояние. Из-за этой особенности их используют в пищевой промышленности в качестве желирующих веществ.
Стенки соседних клеток растений не примыкают друг к другу непосредственно. Между ними находится срединная пластинка, образованная из студнеобразных пектатов магния и кальция.
Соседние клетки растений связаны между собой через плазмодесмы — цитоплазматические мостики, проходящие через отверстия в клеточных стенках и срединных пластинках.
У большинства растительных клеток кроме первичной, после завершения роста и дифференциации, образуется вторичная стенка. Она формируется между цитоплазматической мембраной и первичной оболочкой и состоит из нескольких слоев целлюлозы. При этом фибриллы каждого слоя располагаются под своим углом. Данная структура придает клетке еще большую прочность. Вторичной стенки нет у клеток мягких тканей (например, у мезофилльной ткани листа).
Одревеснение ряда тканей растения связано с так называемой лигнификацией. Вещество лигнин придает стенкам особую прочность и жесткость.
Рассмотрев строение, обратимся к функциям клеточных стенок. У растений нет скелета, однако многие из них достигают огромных размеров, что невозможно без какой-либо внутренней опоры. Ее то совместно и выполняют жесткие оболочки клеток. Итак, главная функция клеточных стенок растений — это обеспечение опоры за счет создания прочного каркаса.
Стенки ограничивают рост клеток и препятствуют их разрыву, не давая в определенных условиях излишкам воды поступать в клетки. Микрофибриллы целлюлозы, ориентируясь определенным образом, определяют направление роста клетки. Так, если волокна преимущественно идут поперек, то рост будет идти вдоль.
У растений есть ткани, выполняющие транспортную функцию. Некоторые из них состоят из мертвых клеток, а функцию транспорта обеспечивают исключительно клеточные стенки.
У некоторых клеток их оболочки служат для хранения запаса питательных веществ.
scienceland.info
Почему клеточная стенка грибов состоит из хитина, а не из целлюлозы, как у растений?
Клеточная стенка — жёсткая оболочка клетки, расположенная снаружи от цитоплазматической мембраны и выполняющая структурные, защитные и транспортные функции. Обнаруживается у большинства бактерий, архей, грибов и растений. Животные и многие простейшие не имеют клеточной стенки. Клеточные стенки грибов состоят из хитина и глюканов. Клеточные стенки высших растений построены в основном из целлюлозы, гемицеллюлозы и пектина. В них существуют отверстия — плазмодесмы, через которые осуществляется контакт соседних клеток и обмен веществами между ними. Растительные клеточные стенки выполняют целый ряд функций: Они обеспечивают жесткость клетки для структурной и механической поддержки, придают форму клетке, направление ее роста и в конечном счете морфологию всему растению. Клеточная стенка также противодействует тургора, т. е. осмотическому давлению, когда дополнительное количество воды поступает в растения. Клеточные стенки защищают против патогенов, проникающих из окружающей среды, и запасают углеводы для растения. Растительные клеточные стенки строятся прежде всего с углеводного полимера целлюлозы. а почему - а потому что так задумано х)
потому что ХГРИБ, и все тут...
touch.otvet.mail.ru
Клеточные стенки грибов
Прокариотическая клетка
Прокариоты — организмы, не обладающие, в отличие от эукариот, оформленным клеточным ядром и другими внутренними мембранными органоидами (за исключением плоских цистерн у фотосинтезирующих видов, например, у цианобактерий). Единственная крупная кольцевая (у некоторых видов — линейная) двухцепочечная молекула ДНК, в которой содержится основная часть генетического материала клетки (так называемый нуклеоид) не образует комплекса с белками-гистонами (так называемого хроматина). К прокариотам относятся бактерии, в том числе цианобактерии (сине-зелёные водоросли), и археи. Потомками прокариотических клеток являются органеллы эукариотических клеток — митохондрии и пластиды. Основное содержимое клетки, заполняющее весь её объём, — вязкая зернистая цитоплазма.
Эукариотическая клетка
Эукариоты — организмы, обладающие, в отличие от прокариот, оформленным клеточным ядром, отграниченным от цитоплазмы ядерной оболочкой. Генетический материал заключён в нескольких линейных двухцепочных молекулах ДНК (в зависимости от вида организмов их число на ядро может колебаться от двух до нескольких сотен), прикреплённых изнутри к мембране клеточного ядра и образующих у подавляющего большинства (кроме динофлагеллят) комплекс с белками-гистонами, называемый хроматином. В клетках эукариот имеется система внутренних мембран, образующих, помимо ядра, ряд других органоидов(эндоплазматическая сеть, аппарат Гольджи и др.). Кроме того, у подавляющего большинства имеются постоянные внутриклеточные симбионты-прокариоты — митохондрии, а у водорослей и растений — также и пластиды.
2) цитоплазма
Цитопла́зма — внутренняя среда живой или умершей клетки, кроме ядра и вакуоли, ограниченная плазматической мембраной. Включает в себя гиалоплазму — основное прозрачное вещество цитоплазмы, находящиеся в ней обязательные клеточные компоненты — органеллы, а также различные непостоянные структуры — включения.
В состав цитоплазмы входят все виды органических и неорганических веществ. В ней присутствуют также нерастворимые отходы обменных процессов и запасные питательные вещества. Основное вещество цитоплазмы — вода.
Цитоплазма постоянно движется, перетекает внутри живой клетки, перемещая вместе с собой различные вещества, включения и органоиды. Это движение называется циклозом. В ней протекают все процессы обмена веществ.
Цитоплазма способна к росту и воспроизведению и при частичном удалении может восстановиться. Однако нормально функционирует цитоплазма только в присутствии ядра. Без него долго существовать цитоплазма не может, так же как и ядро без цитоплазмы.
Важнейшая роль цитоплазмы заключается в объединении всех клеточных структур (компонентов) и обеспечении их химического взаимодействия. Так же цитоплазма поддерживает тургор(объём) клетки, поддержание температуры.
3)Клеточная стенка
Клеточная стенка — жёсткая оболочка клетки, расположенная снаружи от цитоплазматической мембраны и выполняющая структурные, защитные и транспортные функции. Обнаруживается у большинства бактерий, архей, грибов и растений. Животные и многие простейшиене имеют клеточной стенки.
Клеточные стенки прокариот
Клеточные стенки бактерий состоят из пептидогликана (муреина) и бывают двух типов: грамположительного и грамотрицательного. Клеточная стенка грамположительного типа состоит исключительно из толстого слоя пептидогликана, плотно прилегающего к клеточной мембране и пронизанного тейхоевыми илипотейхоевыми кислотами. При грамотрицательном типе слой пептидогликана существенно тоньше, между ним и плазматической мембраной находится периплазматическое пространство, а снаружи клетка окружена ещё одной мембраной, представленной т. н. липополисахаридом и являющаяся пирогенным эндотоксином грамотрицательных бактерий.
Клеточные стенки грибов
Клеточные стенки грибов состоят из хитина и глюканов.
studopedya.ru
1.2 Строение микроскопических грибов
Клеточная стенка. Представляет собой многослойную оболочку из 9...10 слоев различной электронной плотности. Система микрофибрилл, встроенных в аморфный матрикс, формирует скелет клетки. Фибриллы в зависимости от видовой принадлежности могут состоять из целлюлозы, глюкона и хитина. Другие полисахариды, белки, пигменты, липиды служат цементирующими веществами, образующими химические связи с микрофибриллярной частью клеточной стенки. Наличие таких комплексов обеспечивает избирательную проницаемость для одних веществ и блокаду других.
Опорные микрофибриллы клеточной стенки и ее матрикс отличаются по механизму образования и биосинтезу. Образование фибрилл и матрикса происходит несинхронно, в первую очередь регенерируется фибриллярный остов стенки. Биосинтез этих двух частей клеточной стенки осуществляется с участием ферментов.
Процесс образования клеточной стенки происходит двумя способами: новый материал может либо внедряться в стенку поляри- зованно, либо равномерно накладываться по всей ее поверхности. В первом случае происходит образование цилиндрических клеток, во втором — сферических.
Клеточная стенка служит защитным приспособлением и предохраняет грибную клетку от воздействия различных факторов окружающей среды, например осмотическим барьером, обусловливающим избирательную проницаемость для различных веществ. Она придает форму вегетативным клеткам гиф и органов размножения. На поверхности клеточной стенки и цитоплазматической мембраны локализованы ферменты, осуществляющие превращение не усвояемых клеткой (не растворимых в воде) полимеров.
В результате лизиса клеточная стенка грибов может разрушиться под воздействием ферментов, выделяемых другими клетками и образующихся в клетке самого гриба.
Основные компоненты клеточной стенки грибов — хитин, глюканы, белок и жиры. Азотистые и безазотистые полисахариды с жировыми веществами образуют растворимые и нерастворимые комплексы. Основу клеточной стенки составляют 4...6 моносахаров, соотношение которых у различных грибов незначительно варьирует. В состав полисахаридных фракций входят глюкозамин, манноза, глюкоза, ксилоза и др. Следует подчеркнуть, что состав клеточной оболочки различных клеток одного и того же гриба неодинаковый.
Протопласт — содержимое клетки, заключенное в клеточную стенку: Имеет цитоплазматическую мембрану, эндоплазмати- ческий ретикулум, одно или несколько ядер с ядрышками, а также митохондрии, рибосомы с РНК, лизосомы, аппарат Гольджи, вакуоли, пластинчатый комплекс, секреторные гранулы, а также другие структуры и различные включения.
Цитоплазматическая мембрана. Тонкая трехслойная оболочка, располагается непосредственно под клеточной стенкой и отделяет ее от цитоплазмы. Цитоплазматическая мембрана обладает избирательной проницаемостью для веществ, входящих в клетку и выходящих из нее. Цитоплазматическая мембрана содержит до 40 % липидов и до 38 % белков. Различной формы инвагинации и ущемления цитоплазматической мембраны называются мезосомами.
Основное функциональное назначение цитоплазматической мембраны заключается в следующем: осуществление поступления в клетку различных веществ, ферментативная переработка и выделение продуктов метаболизма. Переработанные в цитоплазматической мембране вещества поступают в протопласт клетки и участвуют в обмене веществ.
Эндоплазматический ретикулум. Состоит из пузырьков, канальцев и вакуолей, служащих своеобразным депо питательных веществ.
Митохондрии. Многочисленные подвижные замкнутые образования эллипсовидной формы, с перегородками, покрытые одно- или двухслойной оболочкой. Предполагают, что митохондрии, благодаря собственной ДНК кольцевой структуры, способны к репродукции. Митохондрии окружены мембраной, на которой происходит локализация ферментов: пируватоксидазы, сукциндегидрогеназы, щелочной и кислой фосфатаз, пероксидазы и др. Митохондрии служат генераторами энергии в клетке. В зависимости от условий культивирования и физиологического состояния клетки форма митохондрий и их количество в клетке варьируют.
Рибосомы. Округлые зерна рибонуклеопротеидной природы размером до 200\ принимают участие в синтезе клеточных белков. Количество рибосом значительно отличается у различных видов грибов и зависит от внешних факторов, возраста культуры и др.
Аппарат Гольджи. Представлен группой пузырьков очень мелкого диаметра (0,000 002...О,ООО 01 мкм) или параллельно лежащими дисковидными пластинками. Этот органоид располагается в клетке на участке, свободном от рибосом.
Лизосомы. Производные аппарата Гольджи, размешаются между клеточной оболочкой и цитоплазматической мембраной. Представляют собой зернистые образования, окруженные однослойной липопротеидной мембраной. Содержат фермент, гидролизирующий белок, и выполняют функцию зашиты клеток от неблагоприятного воздействия токсичных веществ экзо- и эндогенного происхождения.
Липосомы. Капельки жировых веществ, окруженные однослойными мембранами.
Ядро. Находится в центре или на полюсах клетки. В грибных клетках могут быть одиночные и множественные ядра. Они отвечают за наследственные функции. Форма ядер округлая или удлиненная. Каждое ядро окружено двухслойной пористой нуклеомембраной с ядрышком из плотных зерен и тонких фибрилл. Ядрышки содержат в составе хромосом ДНК. Через анастомозы ядра могут мигрировать из одной клетки в другую.
Включения. В грибных клетках многочисленные включения: волютин, гликоген, липиды, пигменты, миелоидные образования, соли органических кислот, аминокислоты и др. Считается, что гликоген ответствен за эндогенное дыхание, а волютин служит запасным питательным веществом, участвующим в энергетических процессах.
Следует отметить, что в процессе жизнедеятельности в клетках грибов накапливаются различные продукты метаболизма — антибиотики, ферменты, токсины, витамины и др.
Все многочисленные морфологические элементы микроскопических грибов подразделяют на две группы: мицелий и споры. Они бывают различной формы и размеров. Морфологическое различие спор и мицелия служит важным дифференциальным признаком при определении вида гриба.
Мицелий. Представляет собой узкую круглую трубку, диаметр которой варьирует у микромицетов от одного до нескольких микрон.
Ветвящиеся трубочки — гифы, составляющие мицелий, дифференцируют на более толстые слабо разветвленные и тонкие сильно ветвящиеся. Первые формируют мицелий, главным образом разевающийся на субстрате, вторые — в толще субстрата для поглощения из него питательных веществ. Такая дифференцировка особенно характерна для мицелия некоторых паразитных грибов, но встречается нередко и среди сапрофитных форм. Например, у Rhizopus и некоторых других имеются особые столоны, неветвящиеся и обладающие энергичным ростом.
При обильном ветвлении гифы мицелия, соприкасаясь друг с другом, могут образовывать слияния между клетками, — анастомозы. При наличии большого их количества мицелий приобретает характерный сетчатый вид. Развитие анастомозов наблюдается у различных грибов с многоклеточным мицелием. Благодаря им возможно перемещение клеточного ядра из одной клетки в другую и переход от гаплоидного к диплоидному мицелию. Однако в большинстве случаев они осуществляют вегетативные функции и развиваются у многих форм при недостатке питания. Длина клеток мицелия колеблется от нескольких микрон до десятков и реже сотен микрон.
Мицелий окружен двухконтурной оболочкой, которая у молодых культур более нежная. В перегородках, делящих мицелий на отдельные клетки, имеются поры, через которые в процессе роста переливается цитоплазма, а с ней и питательные вещества. В клетках много различных включений: в старых цитоплазма становится зернистой из-за множества вакуолей. Молодой мицелий состоит из удлиненных прямоугольных клеток, старый — из коротких округлых или многогранных. Мицелий, имеющий перегородки, называется септированным. Однако у некоторых низших грибов мицелий состоит из гиф, лишенных поперечных перегородок, и представляет собой как бы одну, сильно разветвленную гигантскую клетку с многочисленными ядрами и называется несептированным мицелием.
Как же происходит развитие мицелия? Из споры выпячивается ростковая трубочка, которая удлиняется и затем отчленяется перегородкой от средней части, включающей спору. Ростовые трубочки затем еще удлиняются и получают новую перегородку, разделяясь на дистальную, или верхушечную, клетку и проксимальную, или внутреннюю. В дальнейшем верхушечная клетка удлиняется и вновь делится, отделяя вторую, более молодую по сравнению с первой, внутреннюю клетку. Так повторяется и дальше. В этом процессе внутренние клетки только вытягиваются, поперечное деление их происходит редко, но зато из них развиваются боковые ветви. На дистальном конце внутренней клетки образуется боковое выпячивание, принимающее цилиндрическую форму и отделяющееся затем перегородкой от производящей ее клетки. Новая клетка вырастает затем в боковую ветвь, растушую и ветвящуюся таким же образом, как и главная. Благодаря развитию ветвей на протяжении главной гифы они тем старше и сильнее развиты, чем ближе к основанию лежит то их отхождения — акропетальное ветвление.
Развитие несептированного мицелия происходит в общем таким же образом, но без образования поперечных перегородок. Рост происходит на кончиках гиф, где накапливается обильная протоплазма, заполняющая весь просвет, а в более задних частях происходит значительное развитие центральных вакуолей. В однородной среде, например на поверхности питательной желатины, гифы мицелия (как неклеточного, так и многоклеточного) разрастаются равномерно и радиально, так что мицелий имеет форму круга, нарастающего с краев. Центральная часть в нем самая старая, даже иногда отмершая, а периферическая — наиболее молодая.
При общем однообразии развития мицелия, который можно назвать типичным, в отдельных случаях наблюдается ряд специфических черт как макроскопического вида и общего характера роста, так и микроскопического строения. Макроскопический вид мицелия определяют прежде всего воздушные гифы. В одних случаях они формируются на самой поверхности субстрата и отчасти внутри его и тогда мицелий имеет вид плоского, прижатого к субстрату кружка; в других случаях, кроме того, развиваются более или менее обильные гифы, поднимающиеся в воздух и придающие мицелию некоторое сходство, например с куском ваты, возвышающимся над субстратом. Характер роста может быть различным у одного и того же гриба в зависимости от влажности, питания и др. Однако ряд форм грибов имеет специфические особенности, например образование пышного воздушного мицелия — разрушителя древесины.
Цвет мицелия чаше всего бывает снежно-белый, но с возрастом приобретает бурую окраску разных оттенков. Это связано с отложением пигмента в клеточных стенках и реже внутри самой клетки.
Различают мицелий истинный и псевдомицелий. Последний характеризуется тем, что отдельные клетки не связаны друг с другом и не имеют общей оболочки. Вместо истинного ветвления здесь наблюдается древовидное расположение клеток.
Для прикрепления к субстрату и извлечения из него питательных веществ в ходе эволюции у некоторых грибов сформировались специально предназначенные для этого органы: ризоиды и аппрессории, которые учитывают при идентификации грибов. Ризоиды — это корешкообразные, а аппрессории — короткие расширенные, иногда лопастеобразные выросты мицелия.
Грибы, паразитирующие на растениях, иногда формируют специальные ответвления мицелия — гаустории, которые, проникая непосредственно в клетки растений, обеспечивают питание грибов. Для гаусторий типично резкое изменение характера их роста по сравнению с ростом типичного мицелия, что объясняется воздействием протоплазмы живой клетки хозяина.
Склероции, тяжи, ризоморфы и хламидоспоры также являются видоизменениями мицелиального роста.
Склероции представляют собой септированные гифы грибов, образующие особые тела. При формировании склероциев оболочки гиф утолщаются и приобретают темную окраску. Сильно утолщена стенка гиф наружного слоя склероция, внутри же гифы более тонкостенные и обычно не окрашены. Склероции — это защитные приспособительные тела, которые позволяют грибу длительное время сохраняться в окружающей среде и обеспечивают его устойчивость к воздействию различных внешних факторов: температуры, солнечных лучей и др. Зрелые склероции содержат меньше влаги по сравнению с мицелием и много запасных веществ — липидов, гликогена.
Размеры склероциев колеблются от нескольких миллиметров до нескольких десятков сантиметров, а форма бывает самая разнообразная: сферическая, неправильная, в виде прямых или изогнутых рожков и др.
Структура клеток склероциев и механизм их образования различны, однако их формирование происходит путем увеличения ветвления мицелия и септирования гиф. Известны два способа образования склероциев: терминальный — на концах гиф; интеркалярный — в отдельных фрагментах главных гиф.
У многих грибов при развитии плодовых тел и некоторых вегетативных структур образуется ложная ткань — плектенхима (псевдопаренхима). В отличие от настоящей ткани паренхимы, возникающей в результате деления клеток в трех направлениях, плектенхима образуется путем сплетения и срастания. Если она состоит из клеток более или менее изодиаметрических, то ее называют параплектенхимой; если в ней заметно явное гифообразное строение (клетки удлиненной формы), то ее называют прозоплектенхимой.
Мицелиальные тяжи — вегетативная структура линейно агрегированных гиф. Диаметр мицелиальных тяжей зависит от количества гиф, которые концентрируются вокруг центральной основы.
В простейшем случае небольшое количество параллельно идущих гиф склеиваются друг с другом ослизненными наружными оболочками или вступают в более прочное соединение путем фор. мирования многочисленных коротких анастомозов. В других слу. чаях, когда тяжи массивны, их гифы получают определенную дифференцировку. Наружные элементы бывают более тонкими, образуя как бы кору вокруг центрального толстого ствола.
Ризоморфы — более сложные по агрегации гифы, которые отличаются у различных грибов интенсивностью роста центральной гифы, протяженностью боковых ветвлений, а также степенью дифференциации клеток гиф.
Наружные части у ризоморфы обычно темноокрашены и имеют определенное сходство с корнями высших растений. Они широко распространены у грибов с крупными плодовыми телами: у базидиальных, сумчатых и др.
Основное назначение мицелиальных тяжей и ризоморф состоит в обеспечении распространения грибов в субстрате и передвижении по гифам питательных веществ.
Хламидоспоры — это изменения мицелия в зрелых и старых культурах на концах или по его ходу. Основная функция хламидоспор не размножение, а сохранение вида. Форма их обычно круглая, овальная или слегка удлиненная, диаметр превышает диаметр мицелия. У некоторых грибов стенка двухконтурная, поверхность гладкая или шероховатая. Хламидоспоры могут возникать на концах мицелия, тогда они называются терминальными, по ходу мицелия — интерполярными (промежуточными).
В старых культурах часто наблюдают большие скопления хламидоспор причудливой формы, напоминающей четки или ожерелье. Молодые и зрелые хламидоспоры способны прорастать. Старые клетки дегенерируют.
Споры. С помощью спор грибы не только размножаются, но также и распространяются в окружающей среде. Этому способствует высокая устойчивость оболочек спор к воздействию агрессивных факторов. Споры подразделяют на эндоспоры, образующиеся внутри особых вместилищ — спорангиев (сумок), и экзоспоры, располагающиеся на мицелии.
У совершенных грибов споры подразделяют на ооспоры, зигоспоры, аскоспоры, базидиоспоры, эндоспоры, фиалоспоры, хламидоспоры. Споры несовершенных грибов в соответствии с размерами и происхождением также делят на несколько групп. К эндоспорам, образующимся внутри мицелия путем сегментации последнего, относят таллоспоры, включающие в себя артроспоры, хламидоспоры и бластоспоры. Кроме того, для несовершенных грибов характерно образование конидий, макроконидий, алейрий (микроконидий) и гемиспор, считающихся несовершенными конидиями.
Гемиспоры более прочно связаны с мицелием и представляют, собой один или два сегмента, отшнуровывающихся после поперечного деления мицелиальной нити. Форма их цилиндрическая, иногда округлая или многогранная, оболочка двухконтурная.
studfiles.net